പാരമ്പര്യം
മാതാപിതാക്കളിൽ നിന്ന് അവരുടെ സന്തതികളിലേക്ക് സ്വഭാവവിശേഷങ്ങൾ കൈമാറുന്നതാണ് പാരമ്പര്യം അല്ലെങ്കിൽ പൈതൃകം എന്നെല്ലാം അറിയപ്പെടുന്നത്. ഇംഗ്ലീഷിൽ ഇത് ഹെറിഡിറ്റി അല്ലെങ്കിൽ ബയോളജിക്കൽ ഹെറിറ്റൻസ് എന്നെല്ലാം അറിയപ്പെടുന്നു. ഒന്നുകിൽ അലൈംഗിക പുനരുൽപാദനത്തിലൂടെയോ അതല്ലെങ്കിൽ ലൈംഗിക പുനരുൽപാദനത്തിലൂടെയോ, സന്തതി കോശങ്ങൾ അല്ലെങ്കിൽ ജീവികൾ അവരുടെ മാതാപിതാക്കളുടെ ജനിതക വിവരങ്ങൾ സ്വായത്തമാക്കുന്നു. മാതാപിതാക്കളിൽ നിന്നും തുല്യ എണ്ണത്തിലുള്ള ജനതിക ഘടകങ്ങൾ കുട്ടികൾക്ക് ലഭിക്കുന്നു. 23 ക്രോമസോമുകൾ മാതാവിൽ നിന്നും 23 എണ്ണം പിതാവിൽ നിന്നും മനുഷ്യർക്ക് ലഭിക്കുന്നു. ഇങ്ങനെ ലഭിക്കുന്ന 46 ക്രോമസോമുകൾ അഥവാ 23 ജോഡി ക്രോമസോമുകളിൽ അടങ്ങിയിരിക്കുന്ന ഡി.എൻ.എയിൽ മാതാപിതാക്കളുടെ ജനതിക പാരമ്പര്യം വഹിക്കുന്ന ജീനുകൾ കാണപ്പെടുന്നു. പാരമ്പര്യത്തിലൂടെ, വ്യക്തികൾ തമ്മിലുള്ള വ്യതിയാനങ്ങൾ സംഭവിച്ച് പ്രകൃതിനിർദ്ധാരണം വഴി ജീവിവർഗ്ഗങ്ങൾ പരിണമിക്കുകയും ചെയ്യും. ജീവശാസ്ത്രത്തിലെ പാരമ്പര്യത്തെക്കുറിച്ചുള്ള പഠനമാണ് ജനിതകശാസ്ത്രം.
അവലോകനം തിരുത്തുക


പാരമ്പര്യ സ്വഭാവസവിശേഷതകൾ ജീനുകളാൽ നിയന്ത്രിക്കപ്പെടുന്നു. മനുഷ്യരിൽ, കണ്ണുകളുടെ നിറം പാരമ്പര്യ സ്വഭാവത്തിന്റെ ഒരു ഉദാഹരണമാണ്: ഒരു വ്യക്തിക്ക് മാതാപിതാക്കളിൽ ഒരാളിൽ നിന്ന് "തവിട്ട്-കണ്ണിന്റെ സ്വഭാവം" പാരമ്പര്യമായി ലഭിച്ചേക്കാം.[1] ഒരു ജീവിയുടെ ജീനോമിനുള്ളിലെ ജീനുകളുടെ സമ്പൂർണ്ണ സെറ്റിനെ അതിന്റെ ജീനോടൈപ്പ് എന്ന് വിളിക്കുന്നു.[2]
ഒരു ജീവിയുടെ ഘടനയുടെയും പെരുമാറ്റത്തിന്റെയും നിരീക്ഷിക്കാവുന്ന സ്വഭാവസവിശേഷതകളുടെ സമ്പൂർണ്ണ കൂട്ടത്തെ അതിന്റെ ഫിനോടൈപ്പ് എന്ന് വിളിക്കുന്നു. പരിസ്ഥിതിയുമായുള്ള അതിന്റെ ജീനോടൈപ്പിൻ്റെ പ്രതിപ്രവർത്തനത്തിൽ നിന്നാണ് ഈ സ്വഭാവവിശേഷങ്ങൾ ഉണ്ടാകുന്നത്.[3] തൽഫലമായി, ഒരു ജീവിയുടെ ഫിനോടൈപ്പിന്റെ പല വശങ്ങളും പാരമ്പര്യമായി ലഭിക്കുന്നില്ല. ഉദാഹരണത്തിന്, ഒരു വ്യക്തിയുടെ ജനിതകരൂപവും സൂര്യപ്രകാശവും തമ്മിലുള്ള പ്രതിപ്രവർത്തനത്തിൽ നിന്നാണ് സൺടാൻഡ് ചർമ്മം ഉണ്ടാകുന്നത്;[4] അതിനാൽ, സൺടാനുകൾ പാരമ്പര്യമായി കുട്ടികളിലേക്ക് കൈമാറ്റം ചെയ്യപ്പെടുന്നില്ല. എന്നിരുന്നാലും, ചില ആളുകളുടെ ത്വക്കിന് അവരുടെ ജനിതകഘടനയിലെ വ്യത്യാസങ്ങൾ കാരണം മറ്റുള്ളവരെ അപേക്ഷിച്ച് വളരെ എളുപ്പത്തിൽ നിറം കിട്ടുന്നു.[5] ഒരു ശ്രദ്ധേയമായ ഉദാഹരണം ആൽബിനിസത്തിന്റെ പാരമ്പര്യ സ്വഭാവമുള്ള ആളുകളാണ്, അവരുടെ ത്വക്ക് ഒട്ടും നിറമില്ലാത്തതും സൂര്യതാപത്തോട് വളരെ സെൻസിറ്റീവായതുമാണ്.[6]
ഡി.എൻ.എയുടെ പങ്ക് തിരുത്തുക
ജനിതക വിവരങ്ങൾ എൻകോഡ് ചെയ്യുന്ന ഒരു തന്മാത്രയായ ഡിഎൻഎ വഴി പാരമ്പര്യ സ്വഭാവവിശേഷങ്ങൾ ഒരു തലമുറയിൽ നിന്ന് അടുത്തതിലേക്ക് കൈമാറ്റം ചെയ്യപ്പെടുന്നു.[2] പരസ്പരം മാറ്റാവുന്ന നാല് തരം ബേസുകൾ ഉൾക്കൊള്ളുന്ന ഒരു നീണ്ട പോളിമറാണ് ഡിഎൻഎ. ന്യൂക്ലിക് ആസിഡ് സീക്വൻസ് (ഒരു പ്രത്യേക ഡിഎൻഎ തന്മാത്രയ്ക്കൊപ്പമുള്ള ബേസുകളുടെ ക്രമം) ജനിതക വിവരങ്ങൾ വ്യക്തമാക്കുന്നു.[7] ഒരു കോശം മൈറ്റോസിസ് വഴി വിഭജിക്കുന്നതിന് മുമ്പ്, ഡിഎൻഎ പകർത്തപ്പെടുന്നു, അങ്ങനെ തത്ഫലമായുണ്ടാകുന്ന രണ്ട് കോശങ്ങളിൽ ഓരോന്നിനും ഡിഎൻഎ അനുക്രമം ലഭിക്കും. ഒരൊറ്റ പ്രവർത്തന യൂണിറ്റ് വ്യക്തമാക്കുന്ന ഡിഎൻഎ തന്മാത്രയുടെ ഒരു ഭാഗത്തെ ജീൻ എന്ന് വിളിക്കുന്നു. വ്യത്യസ്ത ജീനുകൾക്ക് ബേസുകളുടെ വ്യത്യസ്ത ശ്രേണികളുണ്ട്. കോശങ്ങൾക്കുള്ളിൽ, ഡിഎൻഎയുടെ നീണ്ട സരണികൾ ക്രോമസോമുകൾ എന്ന് വിളിക്കപ്പെടുന്ന ഘടനകൾ ഉണ്ടാക്കുന്നു. ജീനുകൾക്കായി കോഡ് ചെയ്യുന്ന ഡിഎൻഎ സീക്വൻസുകളുടെ സവിശേഷമായ സംയോജനം ഉൾക്കൊള്ളുന്ന ഹോമോലോഗസ് ക്രോമസോമുകളുടെ രൂപത്തിൽ ജീവികൾ അവരുടെ മാതാപിതാക്കളിൽ നിന്ന് ജനിതക വിവരങ്ങൾ പാരമ്പര്യമായി സ്വീകരിക്കുന്നു. ഒരു ക്രോമസോമിനുള്ളിലെ ഡിഎൻഎ ശ്രേണിയുടെ പ്രത്യേക സ്ഥാനം ലോക്കസ് എന്നറിയപ്പെടുന്നു. ഒരു പ്രത്യേക ലോക്കസിലെ ഡിഎൻഎ ക്രമം വ്യക്തികൾക്കിടയിൽ വ്യത്യാസപ്പെടുന്നുവെങ്കിൽ, ഈ ശ്രേണിയുടെ വ്യത്യസ്ത രൂപങ്ങളെ അല്ലീലുകൾ എന്ന് വിളിക്കുന്നു. മ്യൂട്ടേഷനുകൾ വഴി ഡിഎൻഎ സീക്വൻസുകൾ മാറുകയും പുതിയ അല്ലീലുകൾ ഉത്പാദിപ്പിക്കുകയും ചെയ്യാം. ഒരു ജീനിനുള്ളിൽ ഒരു മ്യൂട്ടേഷൻ സംഭവിക്കുകയാണെങ്കിൽ, പുതിയ അല്ലീൽ ജീൻ നിയന്ത്രിക്കുന്ന ട്രെയിറ്റിനെ ബാധിച്ചേക്കാം, ഇത് ജീവിയുടെ ജനിതക സ്വഭാവത്തെ മാറ്റുന്നു.[8]
എന്നിരുന്നാലും, ഒരു അല്ലീലും ഒരു സ്വഭാവവും തമ്മിലുള്ള ഈ ലളിതമായ ഇപാടുകൾ ചില സന്ദർഭങ്ങളിൽ പ്രവർത്തിക്കുന്നുണ്ടെങ്കിലും, മിക്ക സ്വഭാവസവിശേഷതകളും കൂടുതൽ സങ്കീർണ്ണവും ജീവികൾക്കിടയിലും ജീവജാലങ്ങൾക്കിടയിലും ഒന്നിലധികം സംവേദനാത്മക ജീനുകളാൽ നിയന്ത്രിക്കപ്പെടുകയും ചെയ്യുന്നു.[9][10] ജനിതക ശൃംഖലകളിലെ സങ്കീർണ്ണമായ ഇടപെടലുകളും കോശങ്ങൾ തമ്മിലുള്ള ആശയവിനിമയവും ഡവലപ്പ്മെൻ്റൽ പ്ലാസ്റ്റിറ്റിയിലും കനാലൈസേഷനിലുമുള്ള ചില മെക്കാനിക്കുകൾക്ക് അടിവരയിടുന്ന പാരമ്പര്യ വ്യതിയാനങ്ങൾക്ക് കാരണമാകുമെന്ന് ഡവലപ്പ്മെൻ്റൽ ബയോളജിസ്റ്റുകൾ അഭിപ്രായപ്പെടുന്നു. [11]
ഡിഎൻഎ തന്മാത്രയുടെ നേരിട്ടുള്ള ഏജൻസിക്ക് വിശദീകരിക്കാൻ കഴിയാത്ത പാരമ്പര്യ മാറ്റങ്ങളുടെ പ്രധാന ഉദാഹരണങ്ങൾ സമീപകാല കണ്ടെത്തലുകൾ സ്ഥിരീകരിച്ചു. ഈ പ്രതിഭാസങ്ങളെ ജീനുകളിൽ കാര്യകാരണമായോ സ്വതന്ത്രമായോ വികസിക്കുന്ന എപിജെനെറ്റിക് പാരമ്പര്യ വ്യവസ്ഥകളായി തരംതിരിക്കുന്നു. എപിജെനെറ്റിക് പാരമ്പര്യത്തിന്റെ രീതികളെയും സംവിധാനങ്ങളെയും കുറിച്ചുള്ള ശാസ്ത്രീയ ഗവേഷണം ഇപ്പോഴും ശൈശവാവസ്ഥയിലാണ്.[12] ഡിഎൻഎ മെഥിലേഷൻ മാർക്കിംഗ് ക്രോമാറ്റിൻ, സെൽഫ് സസ്റ്റെയിനിങ്ങ് മെറ്റബോളിക് ലൂപ്പുകൾ, ആർഎൻഎ ഇടപെടൽ വഴിയുള്ള ജീൻ സൈലൻസിങ്, പ്രോട്ടീനുകളുടെ ത്രിമാന അനുരൂപീകരണം (പ്രിയോൺ പോലുള്ളവ) എന്നിവ ജൈവ തലത്തിൽ എപിജെനെറ്റിക് പാരമ്പര്യ വ്യവസ്ഥകൾ കണ്ടെത്തിയ മേഖലകളാണ്.[13][14] ഇതിലും വലിയ തോതിലുള്ള ഹെറിറ്റബിലിറ്റി ഉണ്ടാകാം. ഉദാഹരണത്തിന്, നിച്ച് കൺസ്ട്രഷൻ പ്രക്രിയയിലൂടെയുള്ള പാരിസ്ഥിതിക പൈതൃകം നിർവചിക്കുന്നത് അവയുടെ പരിസ്ഥിതിയിലെ ജീവികളുടെ സ്ഥിരമായ ആവർത്തിച്ചുള്ള പതിവ് പ്രവർത്തനങ്ങളാണ്. ഇത് തുടർന്നുള്ള തലമുറകളുടെ തിരഞ്ഞെടുപ്പ് വ്യവസ്ഥയിൽ മാറ്റം വരുത്തുന്നു. പൂർവ്വികരുടെ പാരിസ്ഥിതിക പ്രവർത്തനങ്ങളാൽ സൃഷ്ടിക്കപ്പെട്ട ജീനുകളും പാരിസ്ഥിതിക സവിശേഷതകളും പിൻഗാമികൾക്ക് ലഭിക്കുന്നു.[15] ജീനുകളുടെ നേരിട്ടുള്ള നിയന്ത്രണത്തിലല്ലാത്ത പരിണാമത്തിലെ പാരമ്പര്യത്തിന്റെ മറ്റ് ഉദാഹരണങ്ങളിൽ സാംസ്കാരിക സ്വഭാവങ്ങളുടെ കൈമാറ്റം, ഗ്രൂപ്പ് ഹെറിറ്റബിലിറ്റി, സിംബയോജെനിസിസ് എന്നിവ ഉൾപ്പെടുന്നു.[16][17][18] ജീനിനു മുകളിൽ പ്രവർത്തിക്കുന്ന പൈതൃകതയുടെ ഈ ഉദാഹരണങ്ങൾ മൾട്ടി ലെവൽ അല്ലെങ്കിൽ ഹൈരാർക്കിക്കൽ സെലക്ഷൻ എന്ന തലക്കെട്ടിൽ വിശാലമായി ഉൾക്കൊള്ളിച്ചിരിക്കുന്നു, ഇത് പരിണാമ ശാസ്ത്രത്തിന്റെ ചരിത്രത്തിൽ തീവ്രമായ ചർച്ചയ്ക്ക് വിഷയമാണ്.[17][19]
പരിണാമ സിദ്ധാന്തവുമായുള്ള ബന്ധം തിരുത്തുക
1859-ൽ ചാൾസ് ഡാർവിൻ തന്റെ പരിണാമ സിദ്ധാന്തം അവതരിപ്പിച്ചപ്പോൾ, അതിന്റെ പ്രധാന പ്രശ്നങ്ങളിലൊന്ന് പാരമ്പര്യത്തിന് അടിസ്ഥാനമായ ഒരു സംവിധാനത്തിന്റെ അഭാവമായിരുന്നു.[20] പൈതൃകവും സ്വായത്തമാക്കിയ സ്വഭാവസവിശേഷതകളുടെ (പാൻജെനിസിസ്) പാരമ്പര്യവും മിശ്രണം ചെയ്യുന്നതായി ഡാർവിൻ വിശ്വസിച്ചു. പൈതൃകം മിശ്രണം ചെയ്യുന്നത് ഏതാനും തലമുറകൾക്കുള്ളിൽ ജനസംഖ്യയിലുടനീളം ഏകീകൃതതയിലേക്ക് നയിക്കും, തുടർന്ന് സ്വാഭാവിക തിരഞ്ഞെടുപിൻ്റെ ഭാഗമായി ഒരു ജനസംഖ്യയിൽ നിന്നുള്ള വ്യതിയാനം നീക്കം ചെയ്യും.[21] ഈ ചിന്ത ഓൺ ദി ഒറിജിൻ ഓഫ് സ്പീഷീസിൻ്റെ പിന്നീടുള്ള പതിപ്പുകളിലും അദ്ദേഹത്തിന്റെ പിൽക്കാല ജീവശാസ്ത്ര കൃതികളിലും ഡാർവിൻ ചില ലാമാർക്കിയൻ ആശയങ്ങൾ സ്വീകരിക്കുന്നതിലേക്ക് നയിച്ചു.[22] പാരമ്പര്യത്തോടുള്ള ഡാർവിന്റെ പ്രാഥമിക സമീപനം, മെക്കാനിസങ്ങൾ നിർദ്ദേശിക്കുന്നതിനുപകരം, അത് എങ്ങനെ പ്രവർത്തിക്കുന്നുവെന്ന് രൂപരേഖപ്പെടുത്തുക എന്നതായിരുന്നു (പ്രത്യുൽപാദന സമയത്ത് രക്ഷിതാവിൽ വ്യക്തമായി പ്രകടിപ്പിക്കാത്ത സ്വഭാവവിശേഷങ്ങൾ പാരമ്പര്യമായി ലഭിക്കുമെന്നത് ശ്രദ്ധിക്കുക).
പാരമ്പര്യത്തിന്റെ ബയോമെട്രിക് സ്കൂളിന് ചട്ടക്കൂട് ഉണ്ടാക്കിയ ഡാർവിന്റെ കസിൻ കൂടിയായ ഫ്രാൻസിസ് ഗാൽട്ടൺ ഡാർവിന്റെ പാരമ്പര്യത്തിന്റെ പ്രാരംഭ മാതൃക സ്വീകരിക്കുകയും പിന്നീട് വളരെയധികം പരിഷ്ക്കരിക്കുകയും ചെയ്തു.[23] സമ്പാദിച്ച സ്വഭാവസവിശേഷതകളെ ആശ്രയിച്ചുള്ള ഡാർവിന്റെ പാൻജനസിസ് മോഡലിന്റെ വശങ്ങളെ പിന്തുണയ്ക്കുന്ന തെളിവുകളൊന്നും ഗാൽട്ടൺ കണ്ടെത്തിയില്ല.[24]
1880-കളിൽ ആഗസ്ത് വെയ്സ്മാൻ നിരവധി തലമുറകളിലെ എലികളുടെ വാലുകൾ മുറിച്ചുമാറ്റിയിട്ടും അവയുടെ സന്തതിപരമ്പരയ്ക്ക് വാലുകൾ മുളയ്ക്കുന്നത് തുടരുന്നതായി കണ്ടെത്തിയതിലൂടെ, സ്വായത്തമാക്കിയ സ്വഭാവസവിശേഷതകളുടെ പാരമ്പര്യത്തിന് കാര്യമായ അടിസ്ഥാനമില്ലെന്ന് തെളിയിക്കപ്പെട്ടു.[25]
ചരിത്രം തിരുത്തുക

പുരാതന കാലത്തെ ശാസ്ത്രജ്ഞർക്ക് പാരമ്പര്യത്തെക്കുറിച്ച് പലതരം ആശയങ്ങൾ ഉണ്ടായിരുന്നു. ആൺപൂക്കൾ കാരണം പെൺപൂക്കൾ പാകമാകുന്നുവെന്ന് തിയോഫ്രാസ്റ്റസ് നിർദ്ദേശിച്ചു; [26] "വിത്തുകൾ" വിവിധ ശരീരഭാഗങ്ങളിൽ നിന്ന് ഉത്പാദിപ്പിക്കപ്പെടുകയും ഗർഭധാരണ സമയത്ത് സന്താനങ്ങളിലേക്ക് പകരുകയും ചെയ്യുന്നുവെന്ന് ഹിപ്പോക്രാറ്റസ് അനുമാനിച്ചു;[27] ഗർഭധാരണസമയത്ത് സ്ത്രീ-പുരുഷ ദ്രാവകങ്ങൾ കലരുന്നതായി അരിസ്റ്റോട്ടിൽ കരുതി.[28] ബിസി 458-ൽ എസ്കിലസ്, പുരുഷനെ രക്ഷിതാവായി നിർദ്ദേശിച്ചു.[29]
പാരമ്പര്യത്തെക്കുറിച്ചുള്ള പുരാതന ധാരണകൾ 18-ാം നൂറ്റാണ്ടിൽ വളരെ ചർച്ച ചെയ്യപ്പെട്ട രണ്ട് സിദ്ധാന്തങ്ങളിലേക്ക് മാറി. പാരമ്പര്യത്തെക്കുറിച്ചുള്ള ധാരണയുടെ രണ്ട് വ്യത്യസ്ത വീക്ഷണങ്ങളായിരുന്നു എപ്പിജെനിസിസ് സിദ്ധാന്തവും പ്രീഫോർമേഷൻ സിദ്ധാന്തവും. അരിസ്റ്റോട്ടിൽ നിർദ്ദേശിച്ച എപ്പിജെനിസിസ് സിദ്ധാന്തം, ഭ്രൂണം തുടർച്ചയായി വികസിക്കുന്നുവെന്ന് അവകാശപ്പെട്ടു. മാതാപിതാക്കളുടെ സ്വഭാവസവിശേഷതകൾ ഭ്രൂണത്തിന് അതിന്റെ ജീവിതകാലത്ത് കൈമാറുന്നു. ഈ സിദ്ധാന്തത്തിന്റെ അടിസ്ഥാനം ഇൻഹെറിറ്റൻസ് ഓഫ് അക്വയേഡ് ട്രെയിറ്റ്സ് (നേടിയെടുത്ത സ്വഭാവങ്ങളുടെ അനന്തരാവകാശം) എന്ന സിദ്ധാന്തത്തെ അടിസ്ഥാനമാക്കിയുള്ളതാണ്. നേർവിപരീതമായി, ഡോക്ട്രിൻ ഓഫ് പ്രിഫോർമേഷൻ അവകാശപ്പെടുന്നത് "ഒരേപോലുള്ളവ ഒരേപോലുള്ളവയെ സൃഷ്ടിക്കുന്നു" എന്നതാണ്. വളരെക്കാലം മുമ്പ് സൃഷ്ടിക്കപ്പെട്ടതിനെ വെളിപ്പെടുത്തുന്നതിനുള്ള ഒരു പ്രവർത്തനമാണ് പ്രത്യുൽപാദനമെന്ന് പ്രീഫോർമേഷൻ വീക്ഷണം വിശ്വസിച്ചു. എന്നിരുന്നാലും, പത്തൊൻപതാം നൂറ്റാണ്ടിൽ സെൽ സിദ്ധാന്തത്തിന്റെ സൃഷ്ടിയാണ് ഇത് വിവാദമാക്കിയത്, അവിടെ ജീവന്റെ അടിസ്ഥാന യൂണിറ്റ് കോശമാണ്, അല്ലാതെ ഒരു ജീവിയുടെ മുൻകൂട്ടി തയ്യാറാക്കിയ ചില ഭാഗങ്ങളല്ല. പൈതൃകം മിശ്രണം ചെയ്യുന്നതുൾപ്പെടെയുള്ള വിവിധ പാരമ്പര്യ സംവിധാനങ്ങളും ശരിയായി പരിശോധിക്കപ്പെടുകയോ അളക്കുകയോ ചെയ്യാതെ വിഭാവനം ചെയ്യപ്പെടുകയും പിന്നീട് തർക്കത്തിലാവുകയും ചെയ്തു. എന്നിരുന്നാലും, കൃത്രിമ തിരഞ്ഞെടുപ്പിലൂടെ വളർത്തുമൃഗങ്ങളുടെ ഉല്പ്പാദനവും വിളകളും വികസിപ്പിക്കാൻ ആളുകൾക്ക് കഴിഞ്ഞു. പരിണാമത്തെക്കുറിച്ചുള്ള ആദ്യകാല ലാമാർക്കിയൻ ആശയങ്ങളുടെ ഒരു ഭാഗമാണ് ഇൻഹെറിറ്റൻസ് ഓഫ് അക്വയേഡ് ട്രെയിറ്റ്സ്ഉം.
പതിനെട്ടാം നൂറ്റാണ്ടിൽ, ഡച്ച് മൈക്രോസ്കോപ്പിസ്റ്റ് ആന്റണി വാൻ ലീവൻഹോക്ക് (1632-1723) മനുഷ്യരുടെയും മറ്റ് മൃഗങ്ങളുടെയും ബീജത്തിൽ "അനിമൽകൂൾസ്" കണ്ടെത്തി. [30] ഓരോ ബീജത്തിലും ഒരു "ചെറിയ മനുഷ്യനെ" (ഹോമൺകുലസ്) കണ്ടതായി ചില ശാസ്ത്രജ്ഞർ അനുമാനിച്ചു. ഈ ശാസ്ത്രജ്ഞർ "സ്പെർമിസ്റ്റ്" എന്നറിയപ്പെടുന്ന ഒരു ചിന്താധാര രൂപീകരിച്ചു. ഹോമൺകുലസ് വളർന്നുവരുന്ന ഗർഭപാത്രവും ഗർഭപാത്രത്തിന്റെ ജനനത്തിനു മുമ്പുള്ള സ്വാധീനവും മാത്രമാണ് അടുത്ത തലമുറയുടെ കാര്യത്തിൽ സ്ത്രീയുടെ സംഭാവനയെന്ന് അവർ വാദിച്ചു. സ്പെർമിസ്റ്റ് ചിന്താധാരയെ എതിർക്കുന്ന, ഭാവിയിലെ മനുഷ്യൻ അണ്ഡത്തിലാണെന്നും ബീജം അണ്ഡത്തിന്റെ വളർച്ചയെ ഉത്തേജിപ്പിക്കുന്നുവെന്നും ഓവിസ്റ്റ് ചിന്താധാര പിന്തുടരുന്നവർ വിശ്വസിച്ചു. ഓവിസ്റ്റുകൾ കരുതിയത് സ്ത്രീകളാണ് ആൺകുട്ടികളും പെൺകുട്ടികളും അടങ്ങുന്ന മുട്ടകൾ വഹിക്കുന്നതെന്നും, സന്തതിയുടെ ലിംഗഭേദം ഗർഭധാരണത്തിന് മുമ്പ് തന്നെ നിർണ്ണയിക്കപ്പെട്ടിരുന്നുവെന്നും ആണ്. [31]
1878-ൽ ആൽഫിയൂസ് ഹയാട്ട് കുടുംബത്തിന്റെ പ്രതിഭാസങ്ങളെ (മൂക്കിന്റെ വലിപ്പം, ചെവിയുടെ ആകൃതി മുതലായവ) സംബന്ധിച്ച ഡാറ്റ സമാഹരിച്ച്, രോഗാവസ്ഥകളും അസാധാരണമായ സ്വഭാവസവിശേഷതകളും, പ്രത്യേകിച്ച് പ്രായവുമായി ബന്ധപ്പെട്ട്, പാരമ്പര്യ നിയമങ്ങൾ പഠിക്കാനുള്ള അന്വേഷണത്തിന് നേതൃത്വം നൽകിയപ്പോൾ ഒരു ആദ്യകാല ഗവേഷണ സംരംഭം ഉയർന്നുവന്നു. ചില സ്വഭാവസവിശേഷതകൾ സ്ഥിരമായി പ്രകടിപ്പിക്കുന്നതും മറ്റുള്ളവ വളരെ ക്രമരഹിതമായതും എന്തുകൊണ്ടാണെന്ന് നന്നായി മനസ്സിലാക്കാൻ ഡാറ്റ ടാബുലേറ്റ് ചെയ്യുക എന്നതാണ് പ്രോജക്റ്റുകളിൽ ഒന്ന് ലക്ഷ്യമിടുന്നത്. [32]
ഗ്രിഗർ മെൻഡൽ: ജനിതകശാസ്ത്രത്തിന്റെ പിതാവ് തിരുത്തുക

ജീനുകളുടെ ഇൻഹെറിറ്റൻസ് എന്ന ആശയം ആദ്യം ഉയരുന്നത് മൊറാവിയൻ [33] സന്യാസി ഗ്രിഗർ മെൻഡൽ 1865-ൽ പയറുചെടികളെക്കുറിച്ചുള്ള തന്റെ കൃതി പ്രസിദ്ധീകരിച്ചതോടെയാണ്. എന്നിരുന്നാലും, 1901-ൽ വീണ്ടും കണ്ടെത്തും വരെ അദ്ദേഹത്തിന്റെ കൃതികൾ വ്യാപകമായി അറിയപ്പെട്ടിരുന്നില്ല. മെൻഡലിയൻ ഇൻഹെറിറ്റൻസ് മെൻഡൽ തന്റെ പയറുചെടികളിൽ കണ്ടത് പോലെ വലിയ (ഗുണപരമായ) വ്യത്യാസങ്ങൾക്ക് മാത്രമേ കാരണമായിട്ടുള്ളൂ എന്ന് ആദ്യം അനുമാനിക്കപ്പെട്ടു. ജീനുകളുടെ സങ്കലന (ക്വാണ്ടിറ്റേറ്റീവ്) ഫലത്തെക്കുറിച്ചുള്ള ആശയം ആർഎ ഫിഷറിന്റെ (1918) "The Correlation Between Relatives on the Supposition of Mendelian Inheritance" എന്ന പ്രബന്ധം വരെ യാഥാർത്ഥ്യമായിരുന്നില്ല. മെന്റലിന്റെ പയർ ചെടികളുടെ പ്രദർശനം മെൻഡലിയൻ സ്വഭാവങ്ങളെക്കുറിച്ചുള്ള പഠനത്തിന്റെ അടിത്തറയായി. ഈ സ്വഭാവസവിശേഷതകൾ ഒരൊറ്റ ലോക്കസിൽ കണ്ടെത്താനാകും.
ജനിതകശാസ്ത്രത്തിന്റെയും പാരമ്പര്യത്തിന്റെയും ആധുനിക വികസനം തിരുത്തുക
1930-കളിൽ, ഫിഷറും മറ്റുള്ളവരും ചേർന്ന് നടത്തിയ പ്രവർത്തനങ്ങൾ മെൻഡലിയൻ, ബയോമെട്രിക് സ്കൂളുകളെ ആധുനിക പരിണാമ സമന്വയത്തിലേക്ക് സംയോജിപ്പിച്ചു. ആധുനിക സമന്വയം പരീക്ഷണാത്മക ജനിതകശാസ്ത്രജ്ഞരും പ്രകൃതിശാസ്ത്രജ്ഞരും തമ്മിലുള്ള വിടവ് നികത്തി; രണ്ടും പാലിയന്റോളജിസ്റ്റുകൾക്കിടയിലും ഇത് പ്രസ്താവിക്കുന്നു: [34] [35]
- എല്ലാ പരിണാമ പ്രതിഭാസങ്ങളും അറിയപ്പെടുന്ന ജനിതക സംവിധാനങ്ങളോടും പ്രകൃതിശാസ്ത്രജ്ഞരുടെ നിരീക്ഷണ തെളിവുകളോടും പൊരുത്തപ്പെടുന്ന രീതിയിൽ വിശദീകരിക്കാം.
- പരിണാമം ക്രമാനുഗതമാണ്: ചെറിയ ജനിതക മാറ്റങ്ങൾ, സ്വാഭാവിക തിരഞ്ഞെടുപ്പിന്റെ ക്രമത്തിൽ പുനഃസംയോജനം ചെയ്യപ്പെടുന്നു. ഭൂമിശാസ്ത്രപരമായ വേർതിരിവിലൂടെയും വംശനാശത്തിലൂടെയും ക്രമേണ സ്പീഷീസുകൾ (അല്ലെങ്കിൽ മറ്റ് ടാക്സകൾ) തമ്മിലുള്ള അവ്യക്തത ഉത്ഭവിക്കുന്നതായി വിശദീകരിക്കപ്പെടുന്നു.
- പ്രകൃതി നിർദ്ധാരണം ആണ് മാറ്റത്തിന്റെ പ്രധാന സംവിധാനം;ഇതിൽ ചെറിയ മാറ്റങ്ങൾ പോലും പ്രധാനമാണ്. തിരഞ്ഞെടുക്കാനുള്ള ഒബ്ജക്റ്റ് അതിന്റെ ചുറ്റുമുള്ള പരിതസ്ഥിതിയിലെ ഫിനോടൈപ്പാണ്. തുടക്കത്തിൽ ഡോബ്ജാൻസ്കി ശക്തമായി പിന്തുണച്ചെങ്കിലും ജനിറ്റിക് ഡ്രിഫ്റ്റിന്റെ പങ്ക് അവ്യക്തമാണ്, പാരിസ്ഥിതിക ജനിതകശാസ്ത്രത്തിൽ നിന്നുള്ള ഫലങ്ങൾ ലഭിച്ചതിനാൽ പിന്നീട് അത് തരംതാഴ്ത്തപ്പെട്ടു.
- ജനസംഖ്യാ ചിന്തയുടെ പ്രാഥമികത: സ്വാഭാവിക ജനസംഖ്യയിൽ വഹിക്കുന്ന ജനിതക വൈവിധ്യം പരിണാമത്തിലെ ഒരു പ്രധാന ഘടകമാണ്. കാട്ടിലെ സ്വാഭാവിക തിരഞ്ഞെടുപ്പിന്റെ ശക്തി പ്രതീക്ഷിച്ചതിലും കൂടുതലായിരുന്നു; നിച് അധിനിവേശം പോലെയുള്ള പാരിസ്ഥിതിക ഘടകങ്ങളുടെ സ്വാധീനവും ജീൻ പ്രവാഹത്തിലേക്കുള്ള തടസ്സങ്ങളുടെ പ്രാധാന്യവും എല്ലാം പ്രധാനമാണ്.
ജനസംഖ്യ പ്രത്യുൽപാദനപരമായി ഒറ്റപ്പെട്ടതിന് ശേഷമാണ് സ്പെഷ്യേഷൻ സംഭവിക്കുന്നത് എന്ന ആശയം ഏറെ ചർച്ച ചെയ്യപ്പെട്ടിട്ടുണ്ട്. [36] സസ്യങ്ങളിൽ, സ്പെഷ്യേഷന്റെ ഏത് വീക്ഷണത്തിലും പോളിപ്ലോയിഡി ഉൾപ്പെടുത്തണം. 'പരിണാമം പ്രാഥമികമായി ഒരു തലമുറയ്ക്കും മറ്റൊന്നിനും ഇടയിലുള്ള അല്ലീലുകളുടെ ആവൃത്തിയിലുള്ള മാറ്റങ്ങളാണ്' എന്നതുപോലുള്ള ഫോർമുലേഷനുകൾ പിന്നീട് നിർദ്ദേശിക്കപ്പെട്ടു. വികസന ജീവശാസ്ത്രം (' ഇവോ-ദേവോ ') സംശ്ലേഷണത്തിൽ കാര്യമായ പങ്കുവഹിച്ചില്ല എന്നതാണ് പരമ്പരാഗത വീക്ഷണം, എന്നാൽ സ്റ്റീഫൻ ജെയ് ഗൗൾഡിന്റെ ഗാവിൻ ഡി ബിയറിന്റെ പ്രവർത്തനത്തെക്കുറിച്ചുള്ള ഒരു വിവരണം ഒരു അപവാദമായിരിക്കാമെന്ന് സൂചിപ്പിക്കുന്നു. [37]
സിന്തസിസിന്റെ മിക്കവാറും എല്ലാ വശങ്ങളും ചില സമയങ്ങളിൽ വെല്ലുവിളിക്കപ്പെട്ടിട്ടുണ്ട്. എന്നിരുന്നാലും, പരിണാമ ജീവശാസ്ത്രത്തിൽ സിന്തസിസ് ഒരു വലിയ നാഴികക്കല്ലായിരുന്നു എന്നതിൽ സംശയമില്ല. [38] നിരവധി ആശയക്കുഴപ്പങ്ങൾ നീക്കിയ ഇത് രണ്ടാം ലോകമഹായുദ്ധാനന്തര കാലഘട്ടത്തിൽ ഒരു വലിയ ഗവേഷണത്തെ ഉത്തേജിപ്പിക്കുന്നതിന് നേരിട്ട് ഉത്തരവാദിയായിരുന്നു.
എന്നിരുന്നാലും, ട്രോഫിം ലൈസെങ്കോ, ഇപ്പോൾ സോവിയറ്റ് യൂണിയനിൽ ലിസെൻകോയിസം എന്ന് വിളിക്കപ്പെടുന്ന ഇൻഹെറിറ്റൻസ് ഓഫ് അക്വയേഡ് ട്രെയിറ്റ്സിൽ ഊന്നിയ ലാമാർക്കിയൻ ആശയങ്ങൾ ഊന്നിപ്പറഞ്ഞപ്പോൾ ചില തിരിച്ചു പോക്കുകൾ സംഭവിച്ചു. ഈ പ്രസ്ഥാനം കാർഷിക ഗവേഷണത്തെ ബാധിക്കുകയും 1960-കളിൽ ഭക്ഷ്യക്ഷാമത്തിലേക്ക് നയിക്കുകയും സോവിയറ്റ് യൂണിയനെ സാരമായി ബാധിക്കുകയും ചെയ്തു. [39]
മനുഷ്യരിലും [40] മറ്റ് മൃഗങ്ങളിലും എപിജെനെറ്റിക് മാറ്റങ്ങളുടെ ട്രാൻസ്ജെനറേഷൻ പാരമ്പര്യം ഉണ്ടെന്നതിന് വർദ്ധിച്ചുവരുന്ന തെളിവുകൾ ഉണ്ട്. [41]
സാധാരണ ജനിതക വൈകല്യങ്ങൾ തിരുത്തുക
- ഫ്രാഗിൾ എക്സ് സിൻഡ്രോം
- സിക്കിൾ സെൽ രോഗം
- ഫെനൈൽകെറ്റോണൂറിയ (PKU)
- ഹീമോഫീലിയ
തരങ്ങൾ തിരുത്തുക



ജൈവ പാരമ്പര്യത്തിന്റെ ഒരു രീതിയുടെ വിവരണം മൂന്ന് പ്രധാന വിഭാഗങ്ങൾ ഉൾക്കൊള്ളുന്നു:
- 1. ഉൾപ്പെട്ട ലോക്കസുകളുടെ എണ്ണം
- മോണോജെനെറ്റിക് ("ലളിതമായ" എന്നും അറിയപ്പെടുന്നു) - ഒരു ലോക്കസ്
- ഒലിഗോജെനിക് - കുറച്ച് ലോക്കികൾ
- പോളിജെനെറ്റിക് - ഒരുപാട് ലോക്കികൾ
- 2. ഉൾപ്പെട്ട ക്രോമസോമുകൾ
- ഓട്ടോസോമൽ - ലോക്കികൾ സെക്സ് ക്രോമസോമിൽ സ്ഥിതി ചെയ്യുന്നില്ല
- ഗോണോസോമൽ - ലോക്കുകൾ സെക്സ് ക്രോമസോമിലാണ് സ്ഥിതി ചെയ്യുന്നത്
- എക്സ്-ക്രോമസോമൽ - ലോക്കി എക്സ്- ക്രോമസോമിൽ സ്ഥിതിചെയ്യുന്നു (കൂടുതൽ സാധാരണ കേസ്)
- Y-ക്രോമസോമുകൾ - ലോക്കികൾ Y- ക്രോമസോമിൽ സ്ഥിതിചെയ്യുന്നു
- മൈറ്റോകോൺഡ്രിയൽ - ലോക്കികൾ മൈറ്റോകോൺഡ്രിയൽ ഡിഎൻഎയിലാണ് സ്ഥിതി ചെയ്യുന്നത്
- 3. പരസ്പര ബന്ധ ജീനോടൈപ് - ഫിനോടൈപ്പ്
- ഡോമിനന്റ്
- ഇന്റർമീഡിയറ്റ് (" കോഡോമിനന്റ് " എന്നും അറിയപ്പെടുന്നു)
- റിസസ്സിവ്
- ഓവർഡോമിനന്റ്
- അൺഡർഡോമിനന്റ്
ഈ മൂന്ന് വിഭാഗങ്ങളും മേൽപ്പറഞ്ഞ ക്രമത്തിലുള്ള അനന്തരാവകാശത്തിന്റെ എല്ലാ കൃത്യമായ വിവരണത്തിന്റെയും ഭാഗമാണ്. കൂടാതെ, കൂടുതൽ സ്പെസിഫിക്കേഷനുകൾ ഇനിപ്പറയുന്ന രീതിയിൽ ചേർക്കാം:
- 4. യാദൃശ്ചികവും പാരിസ്ഥിതികവുമായ ഇടപെടലുകൾ
- പെനിട്രൻസ്
- പൂർണ്ണം
- അപൂർണ്ണം (ശതമാനം സംഖ്യ)
- എക്സ്പ്രെസിവിറ്റി
- ഇൻവേരിയബി
- വേരിയബിൾ
- ഹെറിറ്റബിലിറ്റി ( പോളിജെനെറ്റിക്, ചിലപ്പോൾ ഒലിഗോജെനെറ്റിക് ഇൻഹെറിറ്റൻസ് രീതികളിലും)
- മാതൃ അല്ലെങ്കിൽ പിതൃ ഇംപ്രിന്റിങ് പ്രതിഭാസങ്ങൾ (എപിജെനെറ്റിക്സും കാണുക)
- പെനിട്രൻസ്
- 5. ലൈംഗികതയുമായി ബന്ധപ്പെട്ട ഇടപെടലുകൾ
- സെക്സ് ലിങ്ക്ഡ് ഇൻഹെറിറ്റൻസ് ഗോണോസോമൽ ലോക്കി)
- സെക്സ്-ലിമിറ്റഡ് ഫിനോടൈപ്പ് എക്സ്പ്രഷൻ (ഉദാ, ക്രിപ്റ്റോർക്കിസം )
- മാതൃ രേഖയിലൂടെയുള്ള ഇൻഹെറിറ്റൻസ് (മൈറ്റോകോണ്ട്രിയൽ ഡിഎൻഎ ലോക്കിയുടെ കാര്യത്തിൽ)
- പിതൃ രേഖയിലൂടെയുള്ള ഇൻഹെറിറ്റൻസ് ( Y-ക്രോമസോമൽ ലോക്കിയുടെ കാര്യത്തിൽ)
- 6. ലോക്കസ്-ലോക്കസ് ഇടപെടലുകൾ
- മറ്റ് ലോക്കികളുമായുള്ള എപ്പിസ്റ്റാസിസ് (ഉദാ, ഓവർ ഡോമിനൻസ്)
- മറ്റ് സ്ഥലങ്ങളുമായി ജീൻ സംയോജിപ്പിക്കൽ ( കടന്നുപോകുന്നതും കാണുക)
- ഹോമോസൈഗോട്ടസ് മാരക ഘടകങ്ങൾ
- അർദ്ധ-മാരക ഘടകങ്ങൾ
പെഡിഗ്രീ ഡാറ്റയുടെ സ്റ്റാറ്റിസ്റ്റിക്കൽ വിശകലനത്തിലൂടെയാണ് പാരമ്പര്യ രീതിയുടെ നിർണ്ണയവും വിവരണവും പ്രധാനമായും കൈവരിക്കുന്നത്. ഉൾപ്പെട്ടിരിക്കുന്ന സ്ഥലം അറിയാമെങ്കിൽ, മോളിക്യുലാർ ജനിതകത്തിന്റെ രീതികളും ഉപയോഗിക്കാവുന്നതാണ്.
ഡോമിനന്റ്, റിസസീവ് അല്ലീലുകൾ തിരുത്തുക
എല്ലായ്പ്പോഴും ഒരു ജീവിയുടെ രൂപത്തിൽ (ഫിനോടൈപ്പ്) പ്രകടമാകുകയാണെങ്കിൽ, അതിന്റെ ഒരു പകർപ്പെങ്കിലും ഉണ്ടെങ്കിൽ അത് ഡോമിനന്റ് ആണെന്ന് പറയപ്പെടുന്നു. ഉദാഹരണത്തിന്, കടലകളിൽ, പച്ച കായ്കൾക്കുള്ള അല്ലീൽ ആയ G മഞ്ഞ കായ്കളുടെ അല്ലിൽ g യേക്കാൾ ഡോമിനന്റ് ആണ്. അങ്ങനെ GG (ഹോമോസൈഗോട്ട്) അല്ലെങ്കിൽ Gg (ഹെറ്ററോസൈഗോട്ട്) ജോഡി അല്ലീലുകളുള്ള പയർ ചെടികൾക്ക് പച്ച കായ്കൾ ഉണ്ടാകും. മഞ്ഞ കായ്കൾക്കുള്ള അല്ലീൽ റിസസീവ് ആണ്. ഈ അല്ലീലിന്റെ ഫലങ്ങൾ രണ്ട് ക്രോമസോമുകളിലും, gg (ഹോമോസൈഗോട്ട്) ഉള്ളപ്പോൾ മാത്രമേ ദൃശ്യമാകൂ.
-
 എൻസൈമുകളിലെ പാരമ്പര്യ വൈകല്യങ്ങൾ പാരമ്പര്യമായി, എക്സ്-ക്രോമസോമുകളേക്കാൾ കൂടുതൽ നോൺ-എക്സ് ക്രോമസോമുകളുള്ളതിനാൽ സാധാരണയായി ഒരു ഓട്ടോസോമൽ ഫാഷനിലും, ബാധിക്കാത്ത ജീനുകളിൽ നിന്നുള്ള എൻസൈമുകൾ വാഹകരിൽ രോഗലക്ഷണങ്ങൾ തടയാൻ പര്യാപ്തമായതിനാൽ ഒരു റിസസീവ് ഫാഷനിലും ആണ് കാണുന്നത്.
എൻസൈമുകളിലെ പാരമ്പര്യ വൈകല്യങ്ങൾ പാരമ്പര്യമായി, എക്സ്-ക്രോമസോമുകളേക്കാൾ കൂടുതൽ നോൺ-എക്സ് ക്രോമസോമുകളുള്ളതിനാൽ സാധാരണയായി ഒരു ഓട്ടോസോമൽ ഫാഷനിലും, ബാധിക്കാത്ത ജീനുകളിൽ നിന്നുള്ള എൻസൈമുകൾ വാഹകരിൽ രോഗലക്ഷണങ്ങൾ തടയാൻ പര്യാപ്തമായതിനാൽ ഒരു റിസസീവ് ഫാഷനിലും ആണ് കാണുന്നത്. -
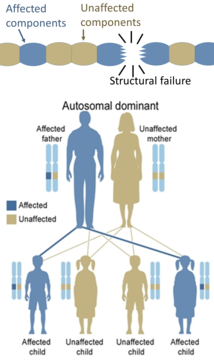 മറുവശത്ത്, ഘടനാപരമായ പ്രോട്ടീനുകളിലെ പാരമ്പര്യ വൈകല്യങ്ങൾ ( ഓസ്റ്റിയോജെനിസിസ് ഇംപെർഫെക്റ്റ, മാർഫാൻ സിൻഡ്രോം, നിരവധി എഹ്ലേഴ്സ്-ഡാൻലോസ് സിൻഡ്രോം എന്നിവ) പൊതുവെ ഓട്ടോസോമൽ ഡൊമിനന്റ് ആണ്, കാരണം മുഴുവൻ ഘടനയും പ്രവർത്തനരഹിതമാക്കാൻ ചില ഘടകങ്ങൾ വികലമായാൽ മതിയാകും. ഇതൊരു ഡോമിനന്റ്-നെഗറ്റീവ് പ്രക്രിയയാണ്, ഇതിൽ ഒരു മ്യൂട്ടേറ്റഡ് ജീൻ ഉൽപ്പന്നം അതേ സെല്ലിനുള്ളിലെ പരിവർത്തനം ചെയ്യപ്പെടാത്ത ജീൻ ഉൽപ്പന്നത്തെ പ്രതികൂലമായി ബാധിക്കുന്നു.
മറുവശത്ത്, ഘടനാപരമായ പ്രോട്ടീനുകളിലെ പാരമ്പര്യ വൈകല്യങ്ങൾ ( ഓസ്റ്റിയോജെനിസിസ് ഇംപെർഫെക്റ്റ, മാർഫാൻ സിൻഡ്രോം, നിരവധി എഹ്ലേഴ്സ്-ഡാൻലോസ് സിൻഡ്രോം എന്നിവ) പൊതുവെ ഓട്ടോസോമൽ ഡൊമിനന്റ് ആണ്, കാരണം മുഴുവൻ ഘടനയും പ്രവർത്തനരഹിതമാക്കാൻ ചില ഘടകങ്ങൾ വികലമായാൽ മതിയാകും. ഇതൊരു ഡോമിനന്റ്-നെഗറ്റീവ് പ്രക്രിയയാണ്, ഇതിൽ ഒരു മ്യൂട്ടേറ്റഡ് ജീൻ ഉൽപ്പന്നം അതേ സെല്ലിനുള്ളിലെ പരിവർത്തനം ചെയ്യപ്പെടാത്ത ജീൻ ഉൽപ്പന്നത്തെ പ്രതികൂലമായി ബാധിക്കുന്നു.


ഇതും കാണുക തിരുത്തുക
അവലംബം തിരുത്തുക
- ↑ Sturm RA; Frudakis TN (2004). "Eye colour: portals into pigmentation genes and ancestry". Trends Genet. 20 (8): 327–332. doi:10.1016/j.tig.2004.06.010. PMID 15262401.
- ↑ 2.0 2.1 Pearson H (2006). "Genetics: what is a gene?". Nature. 441 (7092): 398–401. Bibcode:2006Natur.441..398P. doi:10.1038/441398a. PMID 16724031.
- ↑ Visscher PM; Hill WG; Wray NR (2008). "Heritability in the genomics era – concepts and misconceptions". Nat. Rev. Genet. 9 (4): 255–266. doi:10.1038/nrg2322. PMID 18319743.
- ↑ Shoag J; et al. (Jan 2013). "PGC-1 coactivators regulate MITF and the tanning response". Mol Cell. 49 (1): 145–157. doi:10.1016/j.molcel.2012.10.027. PMC 3753666. PMID 23201126.
- ↑ Pho LN; Leachman SA (Feb 2010). "Genetics of pigmentation and melanoma predisposition". G Ital Dermatol Venereol. 145 (1): 37–45. PMID 20197744. Archived from the original on 2019-03-28. Retrieved 2013-03-26.
- ↑ Oetting WS; Brilliant MH; King RA (1996). "The clinical spectrum of albinism in humans and by action". Molecular Medicine Today. 2 (8): 330–335. doi:10.1016/1357-4310(96)81798-9. PMID 8796918.
- ↑ Griffiths, Anthony, J.F.; Wessler, Susan R.; Carroll, Sean B.; Doebley J (2012). Introduction to Genetic Analysis (10th ed.). New York: W.H. Freeman and Company. p. 3. ISBN 978-1-4292-2943-2.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ↑ Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 978-0-87893-187-3.
- ↑ Phillips PC (2008). "Epistasis – the essential role of gene interactions in the structure and evolution of genetic systems". Nat. Rev. Genet. 9 (11): 855–867. doi:10.1038/nrg2452. PMC 2689140. PMID 18852697.
- ↑ Wu R; Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. 7 (3): 229–237. doi:10.1038/nrg1804. PMID 16485021.
- ↑ Jablonka, E.; Lamb, M.J. (2002). "The changing concept of epigenetics" (PDF). Annals of the New York Academy of Sciences. 981 (1): 82–96. Bibcode:2002NYASA.981...82J. doi:10.1111/j.1749-6632.2002.tb04913.x. PMID 12547675. Archived from the original (PDF) on 2011-05-11.
- ↑ Jablonka, E.; Raz, G. (2009). "Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution" (PDF). The Quarterly Review of Biology. 84 (2): 131–176. doi:10.1086/598822. PMID 19606595. Archived from the original (PDF) on 2011-07-15. Retrieved 2011-02-18.
- ↑ Bossdorf, O.; Arcuri, D.; Richards, C.L.; Pigliucci, M. (2010). "Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana" (PDF). Evolutionary Ecology. 24 (3): 541–553. doi:10.1007/s10682-010-9372-7. Archived from the original (PDF) on 2020-03-01. Retrieved 2019-08-15.
- ↑ Jablonka, E.; Lamb, M. (2005). Evolution in four dimensions: Genetic, epigenetic, behavioural, and symbolic. MIT Press. ISBN 978-0-262-10107-3. Archived from the original on 2021-12-27. Retrieved 2015-06-27.
- ↑ Laland, K.N.; Sterelny, K. (2006). "Perspective: Seven reasons (not) to neglect niche construction". Evolution. 60 (8): 1751–1762. doi:10.1111/j.0014-3820.2006.tb00520.x. PMID 17089961.
- ↑ Chapman, M.J.; Margulis, L. (1998). "Morphogenesis by symbiogenesis" (PDF). International Microbiology. 1 (4): 319–326. PMID 10943381. Archived from the original (PDF) on 2014-08-23.
- ↑ 17.0 17.1 Wilson, D. S.; Wilson, E.O. (2007). "Rethinking the theoretical foundation of sociobiology" (PDF). The Quarterly Review of Biology. 82 (4): 327–348. doi:10.1086/522809. PMID 18217526. Archived from the original (PDF) on 2011-05-11.
- ↑ Bijma, P.; Wade, M.J. (2008). "The joint effects of kin, multilevel selection and indirect genetic effects on response to genetic selection". Journal of Evolutionary Biology. 21 (5): 1175–1188. doi:10.1111/j.1420-9101.2008.01550.x. PMID 18547354.
- ↑ Vrba, E.S.; Gould, S.J. (1986). "The hierarchical expansion of sorting and selection: Sorting and selection cannot be equated" (PDF). Paleobiology. 12 (2): 217–228. doi:10.1017/S0094837300013671. Archived from the original (PDF) on 2016-08-04. Retrieved 2011-02-18.
- ↑ Griffiths, Anthony, J.F.; Wessler, Susan R.; Carroll, Sean B.; Doebley, John (2012). Introduction to Genetic Analysis (10th ed.). New York: W.H. Freeman and Company. p. 14. ISBN 978-1-4292-2943-2.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ↑ Charlesworth, Brian; Charlesworth, Deborah (November 2009). "Darwin and Genetics". Genetics. 183 (3): 757–766. doi:10.1534/genetics.109.109991. PMC 2778973. PMID 19933231. Archived from the original on 2019-04-29. Retrieved 2013-03-26.
- ↑ Bard, Jonathan BL (2011). "The next evolutionary synthesis: from Lamarck and Darwin to genomic variation and systems biology". Cell Communication and Signaling. 9 (30): 30. doi:10.1186/1478-811X-9-30. PMC 3215633. PMID 22053760.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ↑ "Francis Galton (1822-1911)". Science Museum. Archived from the original on January 30, 2016. Retrieved March 26, 2013.
- ↑ Liu Y. (May 2008). "A new perspective on Darwin's Pangenesis". Biol Rev Camb Philos Soc. 83 (2): 141–149. doi:10.1111/j.1469-185X.2008.00036.x. PMID 18429766.
- ↑ Lipton, Bruce H. (2008). The Biology of Belief: Unleashing the Power of Consciousness, Matter and Miracles. Hay House, Inc. pp. 12. ISBN 978-1-4019-2344-0.
- ↑ Negbi, Moshe (Summer 1995). "Male and female in Theophrastus's botanical works". Journal of the History of Biology. 28 (2): 317–332. doi:10.1007/BF01059192.
- ↑ Hipócrates (1981). Hippocratic Treatises: On Generation – Nature of the Child – Diseases Ic. Walter de Gruyter. p. 6. ISBN 978-3-11-007903-6.
- ↑ "Aristotle's Biology – 5.2. From Inquiry to Understanding; from hoti to dioti". Stanford University. Feb 15, 2006. Archived from the original on May 7, 2019. Retrieved March 26, 2013.
- ↑ Eumenides 658-661
- ↑ Snow, Kurt. "Antoni van Leeuwenhoek's Amazing Little "Animalcules"". Leben. Archived from the original on April 24, 2013. Retrieved March 26, 2013.
- ↑ Gottlieb, Gilbert (2001). Individual Development and Evolution: The Genesis of Novel Behavior. Psychology Press. p. 4. ISBN 978-1-4106-0442-2.
- ↑ Scientific American, "Heredity" (in ഇംഗ്ലീഷ്). Munn & Company. 1878-11-30. p. 343. Archived from the original on 2022-05-18. Retrieved 2021-08-06.
- ↑ Henig, Robin Marantz (2001). The Monk in the Garden : The Lost and Found Genius of Gregor Mendel, the Father of Genetics. Houghton Mifflin. ISBN 978-0-395-97765-1.
The article, written by an obscure Moravian monk named Gregor Mendel
- ↑ Mayr & Provine 1998
- ↑ Mayr E. 1982. The growth of biological thought: diversity, evolution & inheritance. Harvard, Cambs. pp. 567 et seq.
- ↑ Palumbi, Stephen R. (1994). "Genetic Divergence, Reproductive Isolation, and Marine Speciation". Annual Review of Ecology and Systematics. 25: 547–572. doi:10.1146/annurev.es.25.110194.002555.
- ↑ Gould S.J. Ontogeny and phylogeny. Harvard 1977. pp. 221–222
- ↑ Handschuh, Stephan; Mitteroecker, Philipp (June 2012). "Evolution – The Extended Synthesis. A research proposal persuasive enough for the majority of evolutionary biologists?". Human Ethology Bulletin. 27 (1–2): 18–21. ISSN 2224-4476.
- ↑ Harper, Peter S. (2017-08-03). "Human genetics in troubled times and places". Hereditas. 155: 7. doi:10.1186/s41065-017-0042-4. ISSN 1601-5223. PMC 5541658. PMID 28794693.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ↑ Szyf, M (2015). "Nongenetic inheritance and transgenerational epigenetics". Trends in Molecular Medicine. 21 (2): 134–144. doi:10.1016/j.molmed.2014.12.004. PMID 25601643.
- ↑ Kishimoto, S; et al. (2017). "Environmental stresses induce transgenerationally inheritable survival advantages via germline-to-soma communication in Caenorhabditis elegans". Nature Communications. 8: 14031. Bibcode:2017NatCo...814031K. doi:10.1038/ncomms14031. PMC 5227915. PMID 28067237.